
抗腫瘤單克降抗體制備策略
19世紀末,人們發現使用相應抗原免疫動物后,獲得含有多克隆抗體的動物抗血清可以用于治療早期的肺炎、白喉、麻疹等傳染病,從而開啟了抗體在醫學中的治療時代。1975年雜交瘤技術的問世,使抗體技術發展進入單克隆抗體的新時代,但鼠源單克隆抗體會產生人抗鼠抗體(human anti-mouse antibody,HAMA)反應,限制了其在臨床上的應用。20世紀70年代,日本醫學家利根川進在基因水平探討了抗體的多樣性形成機制,證實了Ig基因結構,獲得1987年諾貝爾生理學或醫學獎。之后,隨著分子生物學技術的發展,人們開始對抗體進行改造,先后出現了嵌合抗體和人源化抗體,很大程度上解決了鼠抗HAMA反應的問題。20世紀90年代以后,隨著PCR技術、抗體庫技術和轉基因技術的發展,治療性單抗得以實現全人源化改造,目前在臨床上,尤其是在腫瘤、自身免疫性疾病和感染類疾病的治療方面得以廣泛應用。本節將主要探討抗腫瘤單克隆抗體的開發和制備策略。
基于動物免疫的抗體制備策略
雜交瘤抗體技術
1975年德國醫學家Kohler和英國醫學家Milstein開展了一項具有劃時代意義的新技術,他們首次在體外通過將綿羊紅細胞免疫的小鼠脾細胞和小鼠骨髓瘤細胞融合,成功獲得了雜交瘤細胞,經過篩選擴大培養后這種雜交瘤細胞可以產生只針對某一特定的抗原決定簇的抗體,稱為單克隆抗體。這種通過將能無限大量繁殖的腫瘤細胞與能產生抗體的 B細胞融合,以產生既能無限增殖又能產生特異性抗體的技術稱為雜交瘤技術。
雜交瘤技術的主要流程可以概括為:①將免疫動物的脾細胞和無限增殖的瘤細胞融合以得到雜交瘤細胞;②以分泌目標抗體為限制條件,篩選得到的雜交瘤細胞:③擴大培養分泌目標抗體的雜交瘤細胞,使之單克隆化;④運用體外培養或體內誘導法,分離提純以獲得大量的目標單克隆抗體(圖6-1)。
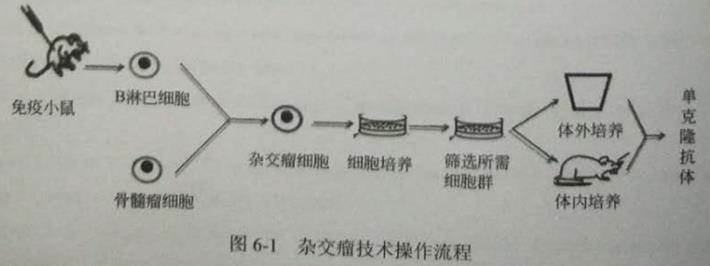
傳統的雜交瘤技術具有生產成本較低、可持續性生產、操作性較好等優點,目前仍然是制備單克隆抗體的主要方法之一。通過雜交瘤技術產生的單克降抗體具有純度高、效價高、特異性好的特點,廣泛應用于生物醫藥、臨床診斷和治療中。但雜交瘤技術也存在一些問題:①是否有合適的骨髄瘤細胞系決定能否有目標克隆抗體產生;②融合形成的雜交瘤細胞也可能低產導致難以分離培養;③因基因不穩定性導致一些難以預料的問題:④因鼠源性抗體的免疫原性,產生HAMA反應,導致機體免疫損傷⑤不能有效激活補體系統且在體內的半衰期較短而起不到良好的免疫效應作用。
人源化抗體
由于單克隆抗體大多數是鼠源性的,在人體內可誘導產生人抗鼠抗體,產生HAMA 反應,限制單克隆抗體在臨床中的應用。隨著基因工程技術的發展及對各類抗體結構、功能和編碼基因認識的深入,可通過將異源抗體中與抗原結合的相關氨基酸結構與人抗體相互組合拼接構成經人源化改造的抗體,以減輕異源性抗體的免疫副作用。
目前對Ig分子結構已經有較清楚的認識,Ig分子的基本結構是一“Y”字形的四肽鏈結構,由兩條相同的重鏈(heavy chain, H)和兩條相同的輕鏈(light chain, L)借助二硫鍵連接起來。其中在多肽鏈的N端,占輕鏈的1/2,重鏈的1/4的氨基酸序列隨抗體的不同而有所變化,稱為可變區(variable region,VR),重鏈和輕鏈的V區分別稱為VH 和VL,各有三個氨基酸排列順序高度可變的區域,稱為高變區(hypervariable region, HVR), 高變區是抗原與抗體特異性結合的位點,其與抗原表位在空間結構上互補,又稱為互補決定簇(complementarity determining region,CDR)。可變區中的四個非髙變區部位氣基酸 排列較為固定,形成骨架結構夾持著CDR,故稱為骨架區(frame work region, FR)。恒 定區(constant region,CR)在多肽鏈的C端,占輕鏈的1/2,重鏈的3/4,其氨基酸數量、 種類、排列順序均較穩定,不同Ig分子的CH長度不一,可以為CH1-3或CH1-4。
對于鼠源性的單克隆抗體的人源化改造研究過程可以分為2個主要的發展階段: ①將鼠源性單克隆抗體的VR和人源性抗體的CR相互組合,形成嵌合抗體:②僅僅保留鼠源性單克隆抗體VR中與抗原結合的CDR,而將FR、CR均改為人源抗體的組成結構, 制備形成的抗體稱為CDR移植抗體或稱改型抗體(圖6-2)。
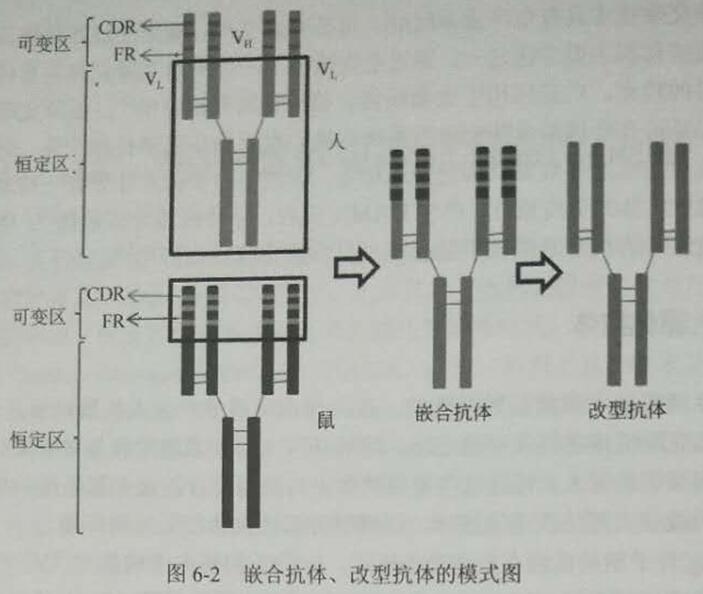
嵌合抗體
嵌合抗體合成的基本原理為:通過利用DNA重組技術,將異源單抗的輕、重鏈可變區基因插入含有人抗體恒定區的表達載體中,轉化哺乳動物細胞表達出嵌合抗體,這樣表達的抗體分子中輕重鏈的V 區是異源的,而C 區是人源的,整個抗體分的近2/3部分都 是人源的。通過這樣的技術合成的抗體,減少了異源性抗體的免疫原性,同時保留了親抗體特異性結合抗原的能力。目前嵌合抗體主一有三種應用形式:嵌合IgG抗體、嵌合Fab抗體、嵌合F(ab')2抗體。嵌合IgG抗體含有人抗體的Fc段能有效的介導細胞免疫治療以及激活補體系統,但鼠源性成分較多,免疫原性大且不易穿透組織,后兩種抗體分子量小、穿透力強,但因不具有人抗體的Fc段而不能直接通過細胞毒作用發揮生物活性,可作為小分子藥物載體或用于診斷試驗。
改型抗體
改型抗體又稱CDR移植抗體,是指抗體的恒定區部分(即CH和CL區)或抗體的全部構成均由人類抗體基因所編碼而產生的抗體,可以明顯減少異源抗體因其免疫原性對人類機體造成的免疫副作用。抗體可變區的CDR直接決定抗體的特異性,是抗體識別和結合抗原的區域。用鼠源性單抗的CDR移植到人源性抗體的可變區替換人源性抗體CDR, 使人源性抗體獲得鼠源性單抗的抗原結合特異性,合成的重組抗體稱為改型抗體,可分為完全CDR移植抗體、部分CDR移植抗體、特異決定區移植抗體三種類型。完全CDR移 植抗體是指將鼠源性單抗的CDR區完全移植到人源性抗體中形成的移植抗體;部分CDR 移植抗體是指由于并不是所有CDR均為抗原抗體特異性結合所必需的,將抗原抗體結合所必需的CDR移植到人源性抗體上以實現減小免疫原性的抗體;特異決定區移植抗體是指將一個CDR中參與抗原識別的特異蛋白分子構成的特異決定區移植替換人源性抗體中相應區域后形成的抗體,其免疫原性有很大程度的降低。然而,雖然抗原與抗體的特異性結合主要與CDR相關,夾持著CDR的支架結構骨架區也可以通過影響CDR的空間構型 來發揮.定的作用,因此重組抗體中鼠源CDR和人源骨架區相嵌的結構可能會導致抗原原有CDR構型的改變,從而導致結合抗原的能力下降。
全人抗體
全人抗體是目前治療性抗體的主要開發趨勢,目前生產全人抗體的技術已達到比較成熟的階段,主耍包括抗體庫技術和基因工程小鼠技術。具體而言,全人抗體是指通過噬菌體展示庫技術、核糖體展示庫技術等抗體庫技術,或者通過將編碼人類抗體的全部基因通過轉基因或轉染色體技術轉移至經過基因工程改造的抗體基因缺失的動物中而表達出人類抗體,來達到抗體全人源化的目的。全人抗體可大幅度地消除異源抗體或部分人源化抗體的免疫原性,避免通過激的免疫副作用,增加抗體應用的安全性,目前已應用于診斷、檢測和臨床治療當中,但也面臨著一些問題,如制備工藝要求較高、不同單抗經過人源化以后悔出現不同程度的與抗原結合能力的下降,一般而言人源化抗體的結合力為原本的33%~50%,在長期、大量、重復應用人源化單抗時,仍然可能會出現HAMA反應。
-
最新內容